自闭症刻板重复行为的病理和可能改善的方向
at 2026.03.24 00:23 ca 自闭症科普 pv 294 by 自闭症科普网
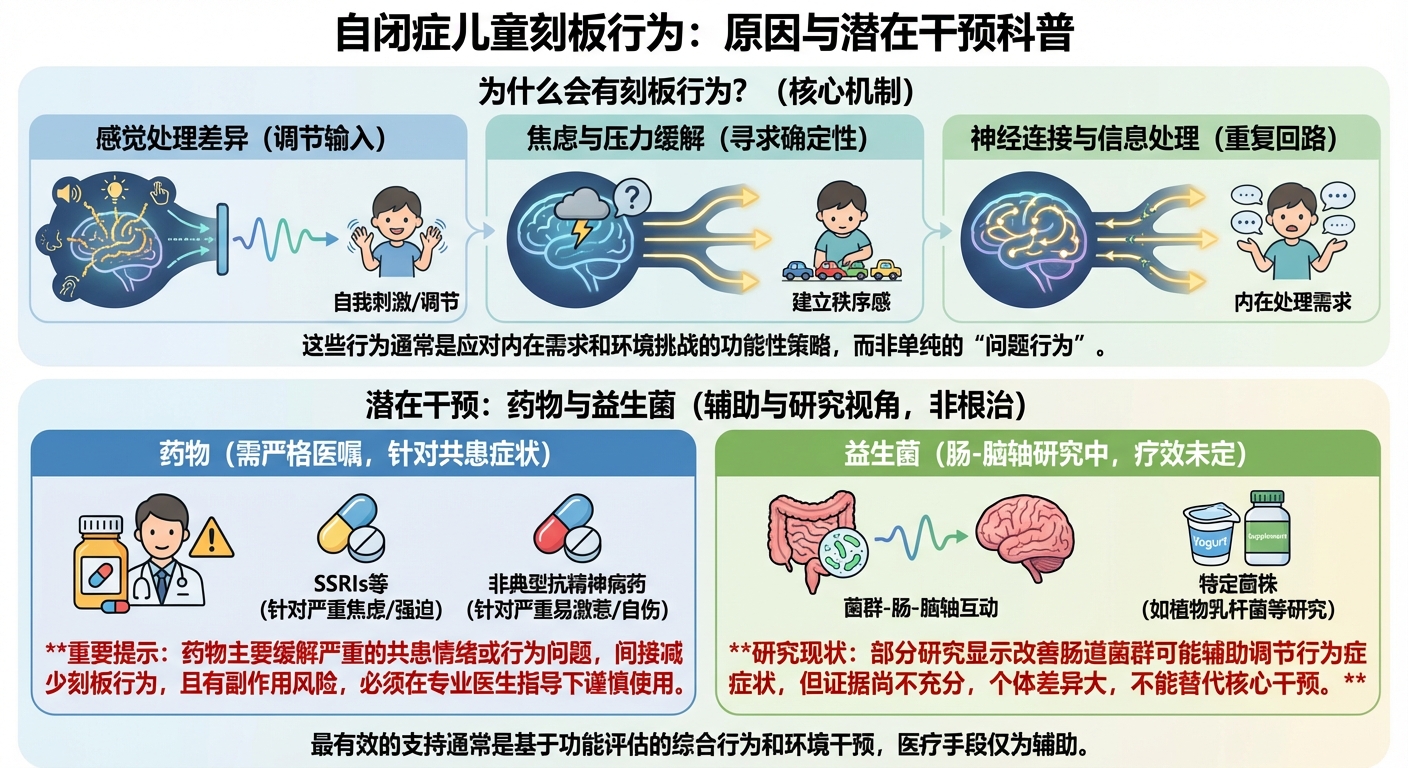
自闭症谱系障碍中刻板重复行为的病理生理机制与多维度干预策略:从神经回路到肠道微生态的深度解析
核心概念的界定与刻板行为的多维地形学特征
自闭症谱系障碍(Autism Spectrum Disorder, ASD)作为一种复杂的神经发育障碍,其临床核心特征主要体现为社交沟通的持续性缺陷以及受限的、重复的行为、兴趣或活动模式。在精神医学领域,刻板行为(Stereotypic Behavior)通常被定义为一类在地形学上表现为固定模式、缺乏明显功能目的或针对性目标的重复性动作。虽然这类行为在演化生物学中可能具有特定的原始功能,但在ASD的临床语境下,它们往往因其高频率、高强度以及在社交场合的不适宜性而成为诊断的关键指标 [1, 2]。
从术语演变的角度看,医学界现已更倾向于使用“限制性与重复性行为”(Restricted and Repetitive Behaviors, RRBs)这一广义范畴,或使用“异常重复行为”(Abnormal Repetitive Behavior, ARB)来涵盖更广泛的临床表现。这类行为具有三个显著的共同特征:极高的重复频率、表现形式的高度不变性以及在当前环境下缺乏明确的目标指向性 [1]。在自闭症儿童的日常生活中,这些行为往往始于幼年期,并随着神经系统的发育而展现出动态的变化趋势。虽然某些刻板行为在儿童成长过程中可能在频率或持续时间上有所下降,但它们通常具有较强的持续性,甚至会演变为更复杂的高级认知仪式 [1, 3]。
为了更精准地理解刻板行为的病理机制,临床研究将其划分为“低级”和“高级”两个集群。低级重复行为主要涉及基础的运动模式,如拍手、身体摇晃、重复性地摆弄物体或自伤行为。这些行为通常表现出明显的节律性,且与感官刺激的自动强化密切相关 [1, 2]。相比之下,高级重复行为则涉及更深层次的认知加工,包括对同一性的极度坚持、对日常例行程序的僵化固守、仪式化的言语或非言语模式,以及高度局限且持久的兴趣。这种分层不仅反映了行为表现的差异,更暗示了大脑不同神经环路参与程度的不同,即从基底神经节的运动反馈到前额叶皮层的认知控制 [1, 3]。
| 刻板行为分类集群 | 核心特征描述 | 典型临床示例 | 潜在功能指向 |
|---|---|---|---|
| 低级运动行为 (Lower-order) | 运动输出的直接重复,多受感觉刺激驱动 | 拍手、转圈、脚趾行走、盯着旋转物体、嗅闻非食物物体 [1, 2] | 感觉寻求、降低生理唤醒度、自动强化 |
| 高级限制行为 (Higher-order) | 认知层面的僵化,表现为对程序和秩序的执着 | 固定的出行路线、特定的排泄仪式、反复询问相同问题、强制排列玩具 [1, 3] | 建立预测感、缓解环境不确定性产生的焦虑 |
| 感觉运动维度 (Sensory-motor) | 对感觉输入的异常反应方式 | 反复开关灯、抓挠皮肤、对特定纹理的舔舐或摩擦 [2, 4] | 调节感觉门控、补偿感觉防御或减退 |
刻板行为对自闭症儿童及其家庭的影响是多维度的。它们不仅会干扰儿童参与正常的教育和社交活动,还往往成为家长压力的主要来源,导致家庭社会适应能力的下降 [1]。然而,必须认识到,这些行为并非单纯的“症状噪音”,它们在个体的神经心理稳态中扮演着自我调节的关键角色 [3]。
刻板行为的神经生物学病因:皮质-纹状体环路与神经递质失衡
深入探究自闭症儿童为何产生刻板行为,首先需要剖析其大脑内部的生物学机制。神经科学研究表明,刻板行为的发生与大脑基底神经节(Basal Ganglia)回路的异常有着直接且深远的联系。基底神经节是一组位于大脑深处的核团,负责运动的启动、抑制以及感觉信息的过滤。在ASD患者中,皮质-纹状体-丘脑-皮质(CSTC)环路的功能性或结构性失衡被认为是刻板行为的核心病理基础 [1]。
基底神经节与运动抑制失效
纹状体(Striatum)作为基底神经节的主要输入端,在调控行为模式中起着“闸门”作用。当纹状体内部的直接通路(促进运动)与间接通路(抑制运动)失去平衡时,大脑对特定运动程序的抑制能力就会减弱,导致某些动作被反复且无意识地释放 [1]。动物模型研究进一步证实,纹状体内多巴胺能神经元的过度活跃或受体敏感性的改变,与重复性运动输出的增加呈显著正相关。此外,解剖学研究发现,ASD儿童的纹状体容量往往表现出异常,这种结构性偏差可能导致感觉信息在传递至皮层进行高级加工前,未能通过基底神经节得到有效的过滤 [1, 5]。
神经递质系统的紊乱
神经递质系统的失衡为刻板行为提供了化学基础。多巴胺(Dopamine)和血清素(Serotonin)是研究最为深入的两种神经递质。多巴胺主要调节奖赏、动机和运动控制,其水平的异常波动往往预示着运动性刻板行为的加剧。血清素则与情绪调节和行为灵活性密切相关,其在突触间隙浓度的不足或受体功能的改变,常导致自闭症个体对环境变化的忍受力降低,从而诱发坚持同一性的高级重复行为 [6, 7]。
此外,兴奋性神经递质谷氨酸与抑制性神经递质 γ-氨基丁酸(GABA)之间的平衡失调也是关键因素。GABA能神经元在大脑中负责抑制不必要的神经放电,若GABA功能减弱,神经系统的整体兴奋性将处于亢进状态,个体更容易表现出过度活跃和感官驱动的刻板动作 [4, 8]。这种化学平衡的紊乱使得ASD儿童的大脑处于一种低抑制状态,使其对内部产生的运动冲动缺乏有效的刹车机制。
感觉加工障碍与刻板行为:一种代偿性的稳态调节机制
自闭症儿童表现出刻板行为的另一个关键原因在于其独特的感觉加工模式。临床数据显示,高达97%的ASD个体存在感觉加工差异,这包括对感觉输入的过度反应(高敏感)、反应不足(低敏感)以及主动的感觉寻求 [4]。
感觉过载与自我保护
对于感觉过度敏感的自闭症儿童来说,普通的物理环境可能充满了“噪音”和“威胁”。明亮的灯光、超市的嘈杂声或特定衣料的触感,都可能引发大脑的感觉过载(Sensory Overload)。在这种状态下,儿童的神经系统会因为接收了过多无法处理的信息而陷入极度痛苦。刻板行为,如反复地蒙住眼睛、捂住耳朵或进行有节奏的身体摇晃,实际上起到了屏蔽外界不可控刺激的作用。通过产生一个高度可预测、自我控制的感官输入(如摇晃产生的本体感觉),儿童能够建立一个暂时的心理庇护所,从而调节过度觉醒的神经系统 [3, 5]。
感觉寻求与神经唤醒
相反,对于感觉反应低下的儿童,他们的神经系统可能长期处于唤醒不足的状态。为了达到正常的心理生理平衡,他们会通过刻板行为来主动寻求强烈的感觉刺激。例如,脚趾行走可以提供更强烈的关节压力反馈,盯着旋转物体或频繁开关灯则能提供强烈的视觉输入。在这种情况下,刻板行为并非病态的干扰,而是大脑为了“自我激活”而采取的一种适应性策略 [2, 5]。
| 感觉加工类型 | 神经生理特征 | 典型的刻板行为表征 | 行为背后的动机 |
|---|---|---|---|
| 感觉过度敏感 | 感觉门控功能受损,无法过滤背景刺激 [5] | 捂耳、闭眼、身体蜷缩、回避特定质地物品 | 降低唤醒水平,逃避痛苦刺激,寻求安全性 [3, 5] |
| 感觉反应低下 | 神经唤醒阈值过高,对普通刺激无感 [5] | 拍手、转圈、碰撞硬物、发出巨大噪音、盯着强光 | 提高唤醒水平,寻求本体感觉和前庭刺激 [2] |
| 感觉调节障碍 | 反应模式在过度与不足之间剧烈波动 [4] | 突发性的情绪爆发伴随激烈的运动性刻板 | 尝试通过节律性动作重新获得神经稳态 [3] |
这种从感觉加工视角出发的解读,促使临床干预从单纯的“行为抑制”转向了环境调控和感觉集成训练。如果这些行为是为了缓解感官压力,那么简单地禁止这些行为可能会导致儿童产生更严重的焦虑或自伤行为。
情绪调节、焦虑与功能性适应:刻板行为的心理学解析
刻板行为不仅仅是生物学缺陷的副产品,它在心理层面承担着重要的情绪调节功能。自闭症儿童由于在社交理解和预测未来事件方面的困难,常年处于高度的焦虑状态中。刻板行为为这种不确定的世界提供了一种“确定性”和“结构感” [3]。
焦虑驱动的自我安抚
研究发现,刻板行为的发生频率与儿童的焦虑水平呈正相关。当环境发生突变(如临时更改行程)或面临社交压力时,儿童会通过重复性的仪式来降低心理压力。这些行为就像是一种“自我安抚”的机制。在神经多样性社区中,这种行为常被称为“Stimming”(自我刺激行为)。对于自闭症个体而言,Stimming是管理情绪过载、焦虑以及处理复杂内部情感状态的重要工具 [3]。
沟通缺失下的替代方案
对于无言语或言语能力受限的自闭症儿童,刻板行为有时承担着非言语沟通的功能。当儿童无法通过语言表达自己的需求(如饥饿、疲劳或社交厌烦)时,他们可能会通过加剧某种刻板动作(如反复敲打头部或快速拍手)来向外界传达其处于不适状态的信息。如果环境无法解读这些信号,行为往往会进一步恶化 [3, 9]。
此外,由于ASD个体在社会互动中往往感到困惑,他们可能会退缩到自己的“刻板世界”中。在这个世界里,一切都是可重复、可预测且受控的。这种对环境的掌控感能够显著降低他们的生存压力,尽管在旁观者看来这些行为是反常的 [2, 3]。
微生物-肠-脑轴在刻板行为中的病理作用
近年来,神经免疫学和肠道微生态学的跨学科研究为理解自闭症刻板行为开辟了全新的视野。微生物-肠-脑轴(Microbiota-Gut-Brain Axis, MGBA)是指肠道菌群、肠道神经系统与中枢神经系统之间通过免疫、内分泌和神经信号传导形成的双向通信网络。越来越多的证据表明,肠道微生态的失衡(Dysbiosis)在ASD的病理过程中起着推波助澜的作用 [10, 11]。
肠道失调与“肠漏”理论
自闭症儿童患胃肠道疾病(如便秘、腹泻、腹痛)的概率是普通儿童的数倍 [10]。这种慢性胃肠道功能紊乱往往伴随着肠道通透性的增加,即所谓的“肠漏”(Leaky Gut)。当肠道屏障受损时,细菌产物如脂多糖(LPS)和各种炎症细胞因子会进入血液循环,并通过受损的血脑屏障(BBB)影响大脑功能。这种全身性的慢性炎症状态会加剧神经炎症,干扰神经突触的修剪和连接,进而影响包括纹状体在内的运动调控区域,使刻板行为更加频繁和剧烈 [11, 12, 13]。
代谢产物对行为的影响
肠道细菌产生的代谢产物,如短链脂肪酸(SCFAs,包括乙酸、丙酸、丁酸),在调节大脑功能中起着关键作用。虽然丁酸对大脑具有一定的保护作用,但异常比例的丙酸在动物实验中已被证实能诱发自闭症样的刻板重复行为。此外,某些肠道病原体(如克雷伯氏菌属、脱硫弧菌属)的过度生长会产生具有神经毒性的代谢产物,干扰神经递质(如多巴胺、血清素)的合成与降解 [11, 14]。这种源自肠道的“生物信号”能够直接跨越距离,影响大脑内部的化学平衡。
| 肠道微生态异常指标 | 潜在的病理机制 | 对刻板行为的潜在影响 |
|---|---|---|
| 微生物多样性降低 (如双歧杆菌减少) [11] | 削弱肠道免疫屏障,增加促炎因子释放 | 维持大脑处于高应激状态,降低行为灵活性 |
| 促炎代谢物 (如脂多糖 LPS) 升高 [11] | 诱发全身性低度炎症,破坏血脑屏障 [12] | 加剧神经炎症,干扰基底神经节对行为的抑制 [12] |
| 神经递质前体代谢紊乱 [13] | 干扰色氨酸代谢,减少中枢血清素合成 | 增加个体的冲动性和对同一性的固执追求 [7] |
| 肠道渗透性增加 (肠漏) [10] | 允许细菌毒素进入循环系统,激活小胶质细胞 | 持续激活大脑免疫防御,加剧刻板行为的频率 |
这种“肠道起源”的观点不仅深化了我们对病因的理解,也为益生菌等生物学干预提供了坚实的理论支撑。
药物干预的临床评估:疗效、风险与管理
针对自闭症儿童的刻板行为,药物治疗通常并非一线选择,但在行为严重影响儿童安全(如自伤)、学习或导致家庭功能崩溃时,药物介入成为必要的辅助手段。目前,FDA尚未批准任何专门针对自闭症刻板行为这一核心症状的药物,但批准了用于治疗关联症状的药物 [15, 16, 17]。
FDA批准的非典型抗精神病药:利培酮与阿立哌唑
利培酮(Risperidone)和阿立哌唑(Aripiprazole)是目前临床上研究最充分、证据等级最高的药物,主要用于治疗ASD相关的“激越”(Irritability),包括攻击性、自伤和剧烈的情绪爆发 [15, 18]。
利培酮 (Risperidone):
作用机制:主要作为多巴胺 D2 受体和血清素 5-HT2A 受体的拮抗剂。
对刻板行为的疗效:多项随机对照试验显示,利培酮在降低异常行为检查表(ABC)中的刻板行为量表得分方面非常有效。它通过降低神经系统的整体唤醒水平,使儿童能够更好地控制其冲动性动作 [7, 15]。
副作用风险:利培酮最显著的副作用是体重显著增加和代谢紊乱。临床观察发现,患儿平均在8周内体重可增加约2.7公斤 [19]。此外,由于其对垂体的作用,可能导致高催乳素血症,引起乳腺发育或月经不调。长期使用的主要风险是迟发性运动障碍(TD),这是一种表现为不自主重复运动的严重副作用 [20, 21]。
阿立哌唑 (Aripiprazole):
作用机制:多巴胺 D2 受体的部分激动剂,被认为具有“多巴胺调节”作用。
疗效特点:在减少激越和多动方面效果显著,起效往往比利培酮更快。虽然它也被认为能减轻某些重复行为,但针对刻板行为的特异性证据稍逊于利培酮 [15]。
安全性 profile:其体重增加的副作用通常比利培酮轻,且不会显著升高催乳素水平。然而,阿立哌唑更容易诱发“静坐不能”(Akathisia),即一种内在的、无法坐定的焦躁感,这在自闭症儿童身上有时会被误认为是多动症的加剧 [15, 18, 19]。
| 药物分类 | 典型药物 | 批准年龄与适应症 | 对刻板行为的获益 | 关键监测指标 |
|---|---|---|---|---|
| 非典型抗精神病药 | 利培酮 () [15] | 5-16岁;激越、攻击、自伤 | 显著降低运动性刻板,改善社交撤退 [7] | 体重、血糖、血脂、催乳素、不自主运动 [19] |
| 非典型抗精神病药 | 阿立哌唑 () [15] | 6-17岁;激越、情绪波动 | 改善行为灵活性,减少破坏性行为 | 静坐不能、疲劳、体重指数 [15] |
| 激动剂 | 胍法辛 / 可乐定 [21] | 非标适应症;伴随多动症 [22] | 降低交感神经张力,缓解焦虑诱发的重复动作 | 血压、心率、镇静状态 [22] |
| 第一代抗精神病药 | 氟哌啶醇 [15] | 非标适应症;严重难治性刻板 | 虽有效但因副作用风险大已少用 [23] | 锥体外系反应 (EPS) [23] |
其他药物的辅助应用
SSRIs (如氟西汀):虽然在成人强迫症中效果显著,但在ASD儿童中的研究结果具有争议。部分儿童在使用后可能出现去抑制现象(激活效应),导致多动和失眠加剧 [16, 24, 25]。
兴奋剂 (如哌甲酯):对于共病ADHD的自闭症儿童,虽然可以改善注意力,但也可能导致激越情绪加重,间接增加刻板行为 [6, 19, 22]。
纳曲酮:作为一种阿片受体拮抗剂,在某些涉及自伤的严重刻板行为中表现出一定潜力,其理论基础是阻断自伤行为带来的“内源性鸦片”快感 [16]。
益生菌及心理益生菌的治疗潜力
鉴于药物治疗的副作用风险,益生菌(Probiotics)作为一种更安全、耐受性更好的干预方式,正受到临床医生的青睐。益生菌通过调节肠道菌群,影响神经递质合成并降低系统性炎症,从而改善行为症状。
植物乳杆菌 PS128 (Lactobacillus plantarum PS128)
PS128是目前针对自闭症行为改善研究最为深入的菌株之一,被归类为“心理益生菌”(Psychobiotics),即通过调节肠道菌群产生精神治疗效果的益生菌 [26, 27]。
临床疗效证据:
在台湾进行的一项为期4周的随机双盲对照试验(RCT)中,71名7-15岁的自闭症男孩在服用PS128后,表现出明显的对立违抗行为减少和焦虑水平下降。更重要的是,家长观察到其刻板行为的频率有所降低 [26, 28]。
另一项开放性研究显示,87%服用PS128的自闭症儿童整体症状得到改善,其中包括显著减少了重复性的身体动作。研究者发现PS128能够调节大脑前额叶皮层和纹状体中的多巴胺及血清素水平 [29, 30]。
作用机制:PS128能够调节下丘脑-垂体-肾上腺(HPA)轴,降低皮质醇水平,从而缓解诱发刻板行为的源头——焦虑。同时,它可能通过迷走神经信号通路,增加脑源性神经营养因子(BDNF)的表达,促进神经的可塑性 [26, 29, 30]。
多菌株混合制剂的协同效应
元分析表明,相比于单菌株,多菌株混合制剂在改善自闭症整体行为症状(ABC总分)方面更具优势。这些合剂通常包含嗜酸乳杆菌、长双歧杆菌和婴儿双歧杆菌 [14, 31, 32]。
改善行为:补充3个月以上的个体在社交互动和重复性言语方面表现出适度的改善 [31, 33]。
缓解胃肠道症状:益生菌能显著改善便秘和腹痛。由于肠道不适往往是刻板行为加剧的诱因(疼痛引发的烦躁),通过消除胃肠痛感,益生菌能够间接降低儿童的情绪不稳定性 [10, 33, 34]。
益生菌干预的实施建议
| 关键要素 | 建议标准 |
|---|---|
| 推荐菌株 | 植物乳杆菌 PS128, 双歧杆菌混合制剂 [26, 35] |
| 建议剂量 | 通常为每日 至 CFU [36, 37] |
| 干预时长 | 至少持续3个月以观察行为变化 [26, 31] |
| 安全性 | 普遍公认为安全 (GRAS),极少数出现轻微胀气 [33, 38] |
尽管益生菌展现了迷人的前景,但它目前更多被视为一种辅助疗法(Adjunctive Therapy),不能完全替代行为干预或必要的药物治疗 [35, 39]。
营养、膳食干预与替代疗法的证据分级
针对刻板行为的生物学管理还包括了膳食改良和微量营养素的补充。
无麸质无酪蛋白 (GFCF) 饮食
GFCF饮食是自闭症家庭中最常尝试的干预手段之一。其理论核心是认为自闭症儿童缺乏分解麸质(小麦)和酪蛋白(乳制品)的酶,生成的阿片类肽会干扰大脑功能 [40, 41]。
临床有效性:一项系统评价指出,GFCF饮食在减少刻板行为(SMD = -0.41)和改善认知方面具有统计学意义。对于那些伴有严重过敏、慢性腹泻或确诊存在食物不耐受的患儿,其效果尤为显著 [40, 42, 43]。
局限与风险:严格执行该饮食极具挑战,且可能导致钙、维生素D和必需氨基酸的摄入不足,从而影响骨骼发育。因此,除非有明确的医学指征,否则不建议盲目进行极端饮食限制 [41, 44]。
维生素D与Omega-3脂肪酸
这两种物质在神经发育和免疫调节中起着核心作用。
维生素D:维生素D受体广泛分布于负责情感加工的大脑区域(如海马体、杏仁核)。研究表明,约57%的ASD儿童存在维生素D缺乏。补充维生素D已被证明能改善行为功能,减少激越感和自伤行为 [8, 45, 46]。
Omega-3 (EPA/DHA):长链多不饱和脂肪酸能改善神经元膜的流动性,并具有强效的抗炎作用。虽然单独补充Omega-3对核心社交症状的改善效果较弱,但它能有效降低多动、冲动和由于感觉过载诱发的重复性动作 [45, 47, 48]。
联合补充维生素D和Omega-3在临床试验中表现出明显的协同效应,能显著降低异常行为检查表中的激越和刻板行为分值 [46, 48]。
综合管理策略与未来展望
理解了刻板行为背后的生理、心理及肠道微生态机制后,临床干预不应再是单一的“纠错”,而应是一个多维度的综合支持体系。
行为干预的一线地位
根据2024-2025年的最新临床指南(如ESCAP和AACAP),应用行为分析(ABA)仍是处理破坏性刻板行为的金标准 [49, 50, 51]。
反应中断与重定向 (RIRD):当儿童陷入有害的刻板动作时,通过立即给予一个简单的、不兼容的任务(如“举起手”来中断“拍手”),能够有效阻断自动强化的路径 [19]。
功能沟通训练 (FCT):通过教导儿童使用图片交换沟通系统(PECS)或手语来表达需求,可以从根本上替代由于挫败感而产生的刻板行为 [9, 50, 52]。
感觉集成与环境支持
由于刻板行为常源于感觉调节障碍,职业疗法(OT)提供的感觉集成训练能够帮助儿童更好地管理感官输入。通过在环境中提供“感觉休息区”(Sensory Breaks)或使用加重背心等工具,可以降低儿童对刻板行为的依赖 [5, 50, 51]。
神经多样性视角下的尊重与容纳
值得注意的是,医学界正经历着一场范式转移。新的干预准则强调,并非所有的刻板行为都需要干预。如果一个重复行为(如轻轻摇晃手指)并不伤害儿童或他人,也不严重干扰学习,它可能是个体在压力社会中生存的必要“减压阀” [52, 53]。强制性地消除这些行为可能会导致儿童发展出更隐蔽、更危险的代偿行为。
未来的研究将继续探索肠脑轴的深层机制,特别是粪菌移植(FMT)等前沿疗法在改善自闭症刻板行为中的长期潜力。目前,FMT在初步试验中显示出比益生菌更持久的症状缓解效果 [11, 38]。
综上所述,自闭症儿童的刻板行为是一个复杂的神经、心理与生物微环境相互作用的结果。有效的干预需要基于详尽的功能评估,综合运用行为策略、感觉支持、针对性的微生态调节(如PS128益生菌)以及在必要时谨慎使用的药物。每个自闭症儿童都是独特的,干预的目标应始终聚焦于提升其生活质量、功能独立性以及在支持性环境中的幸福感。
免责与版权声明
每个“星星的孩子”都是独一无二的,本站分享的科普知识与干预经验仅供学习与交流,不能替代专业医疗机构的评估与诊断。
本站为公益博客,欢迎并鼓励非商业性转载与分享。转载时请注明文章出处,让我们一起传递科学与爱。